Introducción
A menudo, los objetos no son totalmente visibles en la vida cotidiana. Los seres humanos somos capaces de procesar la compleja información visual relacionada con la “incompletitud” porque nuestro entorno visual está compuesto principalmente de objetos opacos que pueden solaparse y ocultarse parcialmente (p. ej. Pepperberg y Nakayama 2016). Los científicos creen que muchas especies no humanas también son capaces de lidiar con la “incompletitud”. Por ejemplo, procesar señales parciales sobre un depredador potencial y reaccionar es más seguro que no reaccionar, incluso cuando surgen falsas alarmas (Figura 1).

Figura 1. En la vida cotidiana los objetos no siempre son plenamente visibles, pero la capacidad de procesar señales parciales, como un posible depredador, puede ser crucial en la vida cotidiana. Crédito de la foto: Tony Alter vía Flickr Creative Common. Haga clic en la imagen para ampliar
Percepción visual amodal
Cuando un objeto parcialmente cubierto todavía se puede observar e identificar fácilmente, como un gato en la hierba o un cuadrado detrás de un círculo, (Figura 2) el proceso es conocido como percepción visual amodal (p. ej. Pepperberg y Nakayama 2016). Mientras que el conocimiento previo y la memoria generalmente juegan un papel en el reconocimiento de los objetos no obstruidos, los procesos perceptivos adicionales (p. ej. una comprensión de la percepción de profundidad, que el círculo está delante del cuadrado) parecen ser necesarios para la percepción visual amodal (Vallortigara 2006 ).
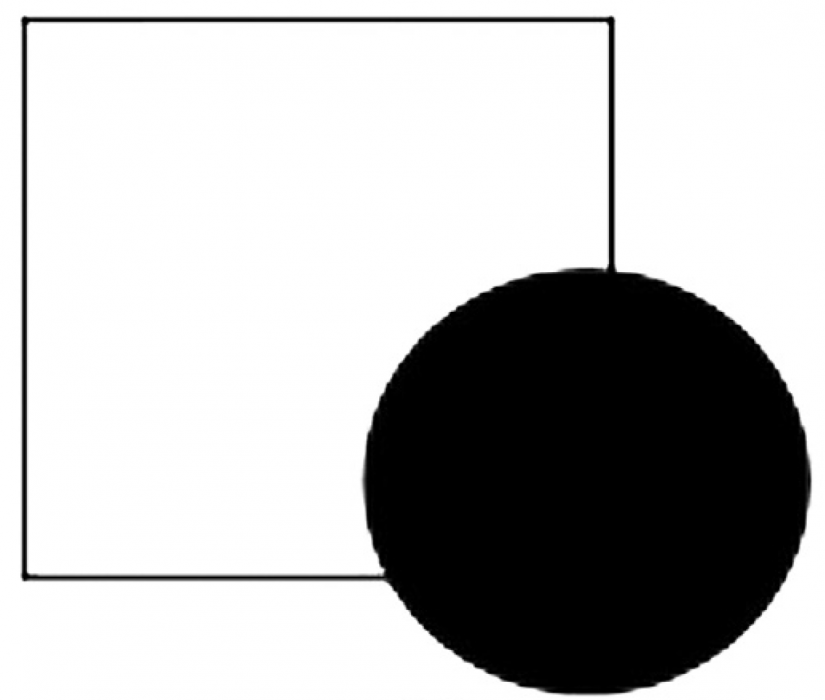
Figura 2. Cuando un objeto parcialmente cubierto todavía se puede observar e identificar fácilmente, el proceso se describe como percepción visual amodal. A pesar que hace falta la esquina inferior derecha, el objeto parcialmente ocluido es rápidamente reconocible como un cuadrado.
Se ha demostrado que varias especies perciben algo sobre los objetos ocluidos, incluyendo los polluelos, minás, urracas y monos (Funk 1996, Pepperberg y Funk 1990, Plowright et al 1998, Vallortigara 2006, revisado en Pepperberg y Nakayama 2016). También se ha evaluado recientemente la evidencia circunstancial de la percepción visual amodal al evaluar la reacción en aves silvestres (Paridae spp.) a modelos ocluidos o amputados de un depredador (Accipiter nisus) colocado junto a un alimentador (Tvardíková y Fuchs 2010). Sin embargo, existen explicaciones alternativas para el comportamiento de estas especies: el diseño del experimento era tal que los sujetos podrían centrarse en aspectos simples de los estímulos sin entender lo que realmente estaban viendo.
No obstante, el concepto de percepción visual amodal parece ser una función ecológicamente válida y omnipresente. Por ejemplo, el reconocimiento de una madre parcialmente ocluida sería útil cuando la cría puede moverse por sí misma para reunirse con ella a fin de restablecer el contacto social. Esta es probablemente la razón por la que el reconocimiento de los objetos parcialmente ocultados emerge tempranamente en los polluelos precociales, pero no en las especies altamente altriciales como los humanos recién nacidos (Vallortigara 2006).
Las palomas y algunos otros animales pueden haber evolucionado con un sistema visual que no completa activamente los estímulos ocluidos, pero los reconoce como dos figuras separadas (Figura 3) (Fujita 2006). Las palomas parecen responder a los estímulos visuales en base a las características locales y visibles y no completan –o incluso perciben– la continuación de la figura detrás del objeto ocluido (Fujita 2006).

Figura 3. Se ha demostrado en estudios que usan una variedad de estímulos y procedimientos, que las palomas no completan figuras parcialmente ocluidas
Plowright et al (1998) encontraron que las palomas perdieron el interés en sus alimentos cuando estos eran colocados detrás de una pantalla y las palomas no los podían ver. Fujita 2001 planteó la hipótesis que estos resultados estaban relacionados con la estrategia nutricional de esta especie. Las palomas son granívoras y debido a que el grano es generalmente abundante, el animal no está obligado a buscar detrás de los obstáculos. La percepción visual amodal puede ser importante para otras especies de aves que se dedican a encontrar y comer una variedad más amplia de alimentos, incluyendo gusanos e insectos que a menudo se esconden bajo las hojas o el suelo y pueden ser sólo parcialmente visibles.
Además, la percepción visual teóricamente requiere más tiempo de procesamiento. Puede ser adaptable para las palomas no completar los estímulos, al menos en las tareas de alimentación comúnmente utilizadas en sus pruebas (Fujita 2006).
Percepción visual modal
Un tipo diferente de función visual implica los contornos subjetivos, o ver figuras que son imaginarias. El término formal para esta función es “percepción visual modal”. El ejemplo más conocido de percepción visual modal es el triángulo de Kanizsa, en el que se puede ver un triángulo limitado por tres figuras “pac-man” colocadas de tal manera que los ángulos abiertos de 60 grados apuntan hacia adentro en la misma región (Figura 4). Además de los contornos subjetivos o ilusorios que crean los bordes de un triángulo, un segundo componente ilusorio consiste en que el triángulo aparece más brillante que la región circundante, aunque esta tenga la misma luminancia física. También se ha demostrado que las abejas y las lechuzas comunes, entre otras especies, perciben algo sobre los contornos subjetivos, pero por las mismas razones descritas para las funciones de percepción visual amodal, los datos son ambiguos (revisado en Pepperberg y Nakayama, 2016).

Figura 4. Los contornos subjetivos o ilusorios son ilusiones visuales que evocan la percepción de un borde sin diferencia de luminancia, color o textura a través de ese borde.
El experimento
Decidimos entonces ver qué pasaría si pudiéramos examinar a un animal de la misma manera que los humanos son examinados: mostrándole estímulos como en las figuras anteriores y simplemente preguntando qué se ve. Encontramos el animal perfecto: Griffin, un loro gris africano macho (Psittacus erithacus) de 16 años de edad, al que previamente se le había enseñado palabras en inglés para los colores y formas de varios objetos sólidos (Figura 5). A diferencia de otros animales, Griffin podría describir vocalmente los elementos de su entorno, ofreciendo una oportunidad única para comparar datos humanos y no humanos (Pepperberg y Nakayama 2016). Sin embargo, Griffin nunca había sido examinado con imágenes y objetos bidimensionales.

Figura 5. Griffin, el loro gris africano (Psittacus erithacus), usó sus habilidades para producir palabras para los colores y formas (llamándolos 1, 2, 3, 4, 6, 8-esquinas). Las formas de ocho esquinas fueron excluidas en los exámenes. Crédito de la foto: Dra. Irene Pepperberg.
El experimento fue diseñado para responder tres preguntas (Pepperberg y Nakayama 2016). Puede un loro gris …
- Reconocer estímulos bidimensionales después de haber sido entrenado en objetos tridimensionales?
- Realizar tareas de identificación utilizando imágenes nuevas para mostrar la presencia de percepción visual modal y amodal?
- Realizar tareas de reconocimiento en objetos en los que no ha sido entrenado y en circunstancias muy diferentes de su entrenamiento?
A pesar de la falta de entrenamiento en objetos bidimensionales como los de las Figuras 1, 4, 6 y 7 y de tener una exposición limitada a objetos ocultos y ninguna exposición a contornos subjetivos, Griffin no tuvo ningún problema con la transición de objetos tridimensionales a bidimensionales. Griffin pudo percibir y etiquetar adecuadamente figuras ocultas y de Kanizsa, demostrando así la percepción amodal y modal (Pepperberg y Nakayama 2016).
Cada uno de los estímulos bidimensionales se sostuvo verticalmente, aproximadamente a 15-20 cm de los ojos de Griffin para atraer su atención (Video 1). Se utilizó la visión monocular porque se desconoce el grado de superposición binocular en el loro gris. El lorito senegalés, una especie que no está muy relacionada con los loros grises, tiene aproximadamente un 30% de superposición binocular (Demery et al 2011).
Video 1. Los estímulos bidimensionales se sostuvieron verticalmente hacia uno de los ojos de Griffin. El experimentador rastreó manualmente la posición del objeto de prueba con respecto a la cabeza del loro para mantener el objeto delante de un ojo.
Para evaluar la percepción visual amodal, los estudiantes de Pepperberg pidieron a Griffin que identificara varios polígonos coloreados que estaban ocluidos por círculos negros (Figura 6). Para la evaluación de la percepción modal, se le mostraron a Griffin figuras de Kanizsa construidas usando “pac-man” negros para formar polígonos regulares en papel coloreado (Pepperberg y Nakayama 2016) (Figura 7).

Figura 6. “¿Qué forma es azul?” Fue una de las preguntas que se usaron para evaluar la percepción visual amodal de Griffin. Nota: Los estímulos mostrados a Griffin no tenían números arábigos.

Figura 7. Se utilizaron nueve figuras de Kanizsa diferentes en este estudio. Se le pidió a Griffin que describiera cuantos vértices estaban presentes en el objeto creado por las figuras “pac-man”. Nota: La mayoría de los experimentos en la literatura no humana usan una figura idéntica para la prueba. Nuevamente, no se incluyeron números en los estímulos presentados a Griffin. Crédito de la foto: Dra. Irene Pepperberg
Los colores utilizados fueron ensayados en un estudio previo para asegurar que Griffin interpretara correctamente la etiqueta de estos colores impresos (Pepperberg et al 2008). Los colores naranja y púrpura fueron ajustados dentro de los rangos apropiados ya que estos colores pueden ser difíciles de identificar por los loros grises africanos (Pepperberg 1994; 2006). La capacidad de las aves psitácidas para ver en el espectro ultravioleta puede hacer que su percepción de color difiera un poco de la de los seres humanos (Pepperberg y Nakayama 2016, Goldsmith y Butler 2005, Carvalho et al 2011, Bowmaker et al 1994).
Los resultados de la evaluación mostraron alta precisión (Tabla 1). Griffin respondió correctamente en la primera parte del experimento, lo que sugirió que él trasladó inmediatamente sus respuestas -sin entrenamiento- de objetos tridimensionales no ocluidos a dibujos de formas ocultas o imaginarias. Griffin respondió a la verdadera forma de las figuras, a pesar que todas estaban parcialmente ocultas o no eran físicamente reales. La mayoría de errores ocurrieron en polígonos y triángulos de una y tres esquinas, los que confundió tanto en las evaluaciones modales como en las amodales (Pepperberg y Nakayama 2016). Sin embargo, estos errores fueron pocos en general y no sugirieron que él se estaba centrando solamente en los ángulos de 60 grados.
| Tabla 1. Resumen de los resultados de la evaluación de Griffin | ||
|---|---|---|
| Parámetro evaluado | Precisión porcentual * | # de ensayos correctos |
| Percepción visual modal | 76 | 29/38 |
| Percepción visual amodal | 70 | 23/33 |
| *El azar se asocia con 20% de precisión | ||
Conclusión
En la experiencia visual de los humanos, cuando un objeto está parcialmente oculto por un obstáculo, no percibimos solamente las piezas o fragmentos de ese objeto. Las partes que son directamente visibles suelen ser suficientes para el reconocimiento de todo el objeto. Los seres humanos también tienden a ver objetos que son creados por ilusiones, como en las figuras Kanizsa. Por lo menos un loro gris africano, Griffin, responde de manera similar y estos resultados —particularmente su transferencia de figuras sólidas tridimensionales a estímulos ocultos e imaginarios bidimensionales, sin ningún entrenamiento— implican que él tiene un concepto de forma sofisticado. Aunque es improbable que esta información sea directamente aplicable cuando se planea el enriquecimiento ambiental, nos recuerda la inteligencia de estas aves y la necesidad de tal estimulación en su ambiente.
**Inicia sesión para ver las referencias**
Referencias
Referencias
Bowmaker JK, Heath LA, Das D, Hunt DM. Spectral sensitivity and opsin structure of avian rod and cone visual pigments. Invest Ophthalmol Vis Sci 35:1708, 1994.
Carvalho LS, Knott B, Berg ML, et al. Ultraviolet-sensitive vision in long-lived birds. Proc Royal Society B 278:107-114, 2011. Disponible en http://rspb.royalsocietypublishing.org/content/278/1702/107. Consultado Julio 28, 2016.
Fujita K. Seeing what is not there: Illusion, completion, and spatiotemporal boundary formation in comparative perspective. In: Wasserman EA, Zentall TR (eds). Comparative Cognition: Experimental Explorations of Animal Intelligence. New York, NY: Oxford University Press; 2006: 37, 42-50.
Fujita K. Perceptual completion in rhesus monkeys (Macaca mulatta) and pigeons (Columba livia). Perception and Psychophysics 63:115-125, 2001.
Funk MS. Development of object permanence in the New Zealand parakeet (Cyanoramphus auriceps). Animal Learning and Behavior 24:375-383, 1996.
Goldsmith T, Butler BK. Color vision of the budgerigar (Melopsittacus undulates): Hue matches, tetrachromacy, and intensity discrimination. J Comp Physiol 191:933-951, 2005. Disponible en http://www.2ndchance.info/tameparrot-Goldsmith2005.pdf. Consultado Julio 28, 2016.
Pepperberg IM. Addition by a Grey parrot (Psittacus erithacus), including absence of quantity. J Comp Psychol 120:1-11, 2006.
Pepperberg IM. The Alex Studies; Cognitive and Communicative Abilities of Grey Parrots. Cambridge, MA: Harvard; 1999.
Pepperberg IM. Evidence for numerical competence in an African Grey parrot (Psittacus erithacus). J Comp Psychol 106:295-305, 1994.
Pepperberg IM, Funk MS. Object permanence in four species of psittacine birds: An African Grey parrot (Psittacus erithacus), an Illiger mini maaw (Ara maracana), a parakeet (Melopsittacus undulatus), and a cockatiel (Nymphicus hollandicus). Animal Learning and Behavior 18:97-108, 1990
Pepperberg IM, Nakayama K. Robust representation of shape in a Grey parrot (Psittacus erithacus). Cognition 153:146-160, 2016.
Pepperberg IM, Vicinay J, Cavanagh P. The Muller-Lyer illusion is processed by a Grey parrot (Psittacus erithacus). Perception 37:765-781, 2008.
Plowright CMS, Reid S, Kilian T. Finding hidden food: Behavior on visible displacement tasks by mynahs (Gracula religiosa) and pigeons (Columba livia). J Comp Psychology 112:13-25, 1998.
Tvardíková K, Fuchs R. Tits use amodal completion in predator recognition: A field experiment. Animal Cognition 13:609-615, 2010.
Vallortigara G. The cognitive chicken: Visual and spatial cognition in a nonmammalian brain. In: Wasserman EA, Zentall TR (eds). Comparative Cognition: Experimental Explorations of Animal Intelligence. New York, NY: Oxford University Press; 2006: 53-70.