Introduction
Les objets sont souvent incomplètement visibles dans la vie du quotidien. Les êtres humains sont capables de traiter une information visuelle complexe reliée à l’ « incomplèteté » car notre environnement visuel est principalement composé d’objets opaques qui peuvent se superposés et se masquer partiellement les uns les autres (par exemple Pepperberg et Nakayama 2016). Les scientifiques pensent que de nombreuses espèces non humaines sont également en capacité de gérer cette incomplèteté. Par exemple, interpréter des indices partiels concernant un prédateur potentiel et réagir en conséquence est plus sécuritaire que de ne pas le faire, même si une fausse alarme est déclenchée (Fig 1).

Figure 1. Les objets ne sont pas toujours visibles dans leur intégralité au quotidien, pourtant la capacité de traiter des indices partiels, tels qu’un prédateur potentiel peut être cruciale dans la vie de tous les jours. Crédit photographique : Tony Alter via Flickr Creative Common.
Le remplissage amodal
Lorsqu’un objet partiellement caché est toujours facilement vu et identifié, comme un chat dans l’herbe ou un carré derrière un cercle (Fig 2), le processus impliqué est un exemple de remplissage amodal (par exemple, Pepperberg et Nakayama 2016). Bien que les connaissances antérieures et la mémoire jouent généralement un rôle dans la reconnaissance d’objets cachés, des processus de perception additionnels (par exemple, une compréhension de la perception de profondeur, que le cercle est en face du carré) semble être requis pour le remplissage amodal (Vallortigara 2006).
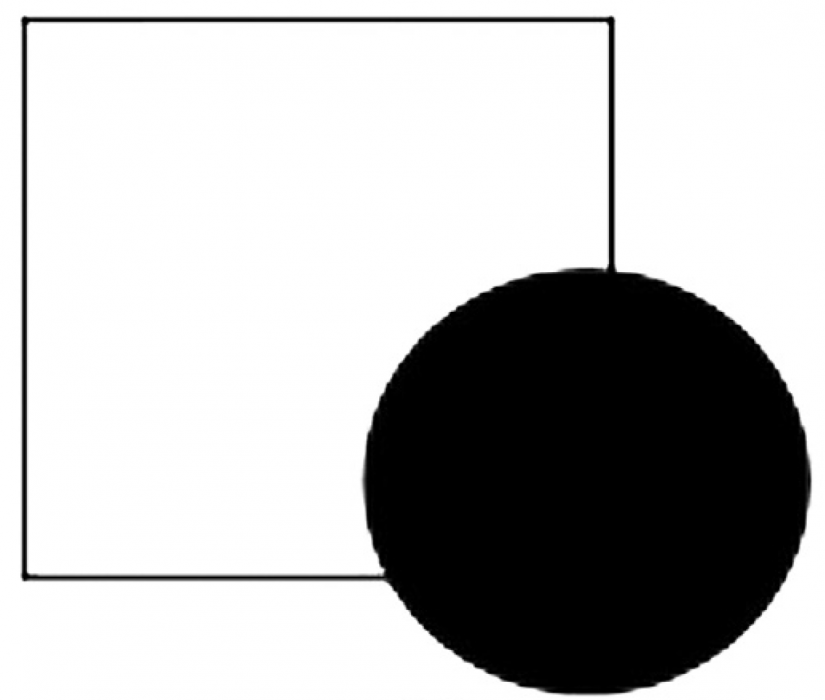
Figure 2. On parle de remplissage amodal lorsqu’un objet partiellement masqué est toujours facilement vu et identifié. Malgré le coin inférieur droit manquant, l’objet partiellement masqué est rapidement reconnu comme étant un carré.
Il a été prouvé qu’un certain nombre d’espèce, incluant les poussins, les mainates, les pies et les singes, avaient une perception des objets masqués (Funk 1996, Pepperberg and Funk 1990, Plowright et al 1998, Vallortigara 2006; revue dans Pepperberg and Nakayama 2016). Des preuves circonstancielles de remplissage amodal ont été récemment évaluée en testant la réaction d’oiseaux sauvages (Paridae spp.) à des modèles cachés ou amputés de prédateurs (Accipiter nisus), fixés sur une mangeoire (Tvardíková and Fuchs 2010). Toutefois, des explications alternatives existent pour comprendre le comportement de ces espèces : la conception de leur tâche était telle que les sujets pouvaient ne se concentrer que sur des aspects simples des stimuli sans comprendre ce qu’ils étaient vraiment en train de regarder, par exemple, en associant l’angle de 90° dans le coin supérieur gauche de la figure 1 à celui d’une autre forme testée sans nécessairement reconnaître que la figure est en fait un carré masqué, ou dans le cas des oiseaux sauvages sans avoir l’expérience d’un rapace partiellement caché qui s’envole pour une attaque.
Néanmoins, le concept de remplissage amodal semble être une tâche écologique valide et ubiquitaire. Par exemple, la reconnaissance d’une mère partiellement cachée parait utile chez des individus capables de se déplacer seuls, de sorte à ce qu’ils puissent la rejoindre et réinstaurer un contact social. Ceci est probablement la raison pour laquelle la reconnaissance d’objet partiellement caché se développe précocément chez les oiseaux précociaux mais pas chez les espèces hautement altriciales comme le nouveau-né humain. (Vallortigara 2006)
Les pigeons, et certains autres animaux, ont pu évolué avec un système visuel qui ne complète pas de façon active des stimuli masqués, mais les reconnaissent comme deux figures séparées (par exemple, Fig 3)(Fujita 2006). Les pigeons semblent répondre à des stimuli visuels sur la base de caractéristiques visibles et locales, et échouent à compléter – ou même à percevoir – la continuité d’une figure en arrière de l’objet masqué (Fujita 2006).

Figure 3. Il a été prouvé que les pigeons ne complètent pas des figures partiellement cachées dans des études utilisant une variété de procédures et de stimuli.
Plowright et al (1998) ont trouvé que les pigeons perdaient leur intérêt pour la nourriture lorsque celle-ci était invisible derrière un écran. Fujita 2001 a supposé que ceci était relié à la stratégie nutritionnelle chez cette espèce. Les pigeons sont des consommateurs de grains et, du fait que les grains sont habituellement abondants, cet animal n’a pas besoin de le chercher sous des obstacles. Le remplissage amodal pourrait être important pour d’autres espèces d’oiseaux qui sont impliqués dans la recherche et la consommation d’une plus grande variété d’aliments, incluant les vers et les insectes, qui se cachent souvent sous les feuilles ou la terre et ne peuvent être que partiellement visibles.
De plus, le remplissage demande théoriquement plus de temps de traitement de l’information. Il peut être adaptatif chez les pigeons de ne pas compléter les stimuli, du moins pas pour les tâches d’alimentation communément utilisées lors des tests (Fujita 2006).
Le remplissage modal
Un type différent de tâche visuel implique les contours subjectifs, c’est-à-dire le fait de voire des figures qui sont en fait imaginaires. Le terme formel est « remplissage modal ». L’exemple le plus connu de remplissage modal est le triangle de Kanizsa, dans lequel une personne peut voire un triangle délimité par trois figures en « pac-man » positionnées de telle sorte que les angles ouverts de 60 degrés pointent tous à l’intérieur vers la même région (Fig 4). En plus des contours subjectifs ou illusoires qui créent les bords d’un triangle, un deuxième composant illusoire est représenté par le fait que le triangle apparait plus clair que la région l’entourant, bien qu’ayant la même luminosité physique. Il a également été prouvé que les abeilles et les effraies des clochers, parmi d’autres espèces, avaient une perception des contours subjectifs, mais pour les mêmes raisons décrites pour la réalisation d’un remplissage amodal, les données sont ambiguës (revue dans Pepperberg et Nakayama 2016).

Figure 4. Les contours subjectifs ou illusoires sont des illusions d’optique qui évoquent la perception d’un trait sans différence de luminosité, de couleur ou de texture de part et d’autre de ce trait.
L’expérimentation
C’est dans ce contexte que nous avons décidé de voire ce qui se passerait s’il était possible de tester un non humain, exactement de la même manière que les humains sont testés : en présentant des stimuli comme dans les figures ci-dessus et en demandant simplement ce qui était vu. Nous avions le sujet parfait : Griffin, un gris du Gabon (Psittacus erithacus) mâle âgé de 16 ans, qui avait au préalable appris les mots en anglais pour les couleurs et les formes de plusieurs objets solides (Fig 5). Au contraire de nombreux sujets animaux, Griffin pouvait décrire vocalement les objets dans son environnement, offrant une opportunité unique de comparer les données humaines et non humaines (Pepperberg et Nakayama 2016). Il n’avait, toutefois, jamais été testé sur des images en deux dimensions des objets.

Figure 5. Griffin le gris du Gabon (Psittacus erithacus) a utilisé ces capacités à caractériser les couleurs et les formes (en les dénommant 1,2,3,4,6,8-angles). Les formes à 8 angles étaient exclues de l’expérience. Crédit photographique : Dr Irene Pepperberg
L’expérimentation était conçue de sorte à pouvoir répondre à trois questions (Pepperberg et Nakayama 2016). Est-ce qu’un gris du Gabon peut…
- Reconnaître des stimuli en deux dimensions après avoir été entrainé sur des objets en trois dimensions ?
- Réaliser des tâches d’identification en utilisant des nouvelles images pour montrer l’existence d’un remplissage modal et amodal
- Reconnaître des objets pour lesquels il n’avait pas été entrainé et dans des conditions très différentes de son entraînement.
Malgré l’absence d’entrainement sur des objets à deux dimensions tels que ceux des Figures 1,4, 6 et 7, ainsi qu’une exposition limitée à des objets masqués et aucune exposition à des contours subjectifs, Griffin a réalisé un transfert immédiat de son apprentissage sur les objets en trois dimensions à des tests sur des stimulis à deux dimension. Griffin pouvait percevoir et caractériser de façon appropriée des figures masqués et de Kanizsa, démontrant ainsi à la fois un processus de remplissage amodal et modal (Pepperberg et Nakayama 2016).
Chacun des stimuli en 2D était maintenu verticalement, à approximativement 15-20 cm des yeux de Griffin pour attirer son attention (Vidéo 1). La vision monoculaire était utilisée car l’étendue de la superposition binoculaire chez les Gris du Gabon est inconnue. Chez le youyou du Sénégal, une espèce qui n’est pas très proche des Gris du Gabon, la superposition binoculaire est d’environ 30% (Demery et al 2011).
Video 1. Des stimuli bidimensionnels étaient maintenus verticalement vers l’un des yeux de Griffin. L’expérimentateur orientait manuellement la position de l’objet testé par rapport à la tête du perroquet pour maintenir une présentation en face d’un seul œil.
Afin de tester le remplissage amodal, les étudiants de Pepperberg demandaient à Griffin de caractériser différents polygônes colorés masqués par des cercles noires (Fig 6). Pour l’évaluation du remplissage modal, des figures de Kanizsa construites en utilisant des « pac-men » noirs formant des polygones réguliers sur du papier coloré étaient présentées à Griffin (Pepperberg y Nakayama 2016) (Fig 7)

Figure 6. “Quelle est la forme bleue ?” était l’une des questions utilisées afin de tester Griffin pour le remplissage amodal. Note : Aucun chiffre arabe ne se trouvait sur les stimuli de Griffin

Figure 7. Neuf figures de Kanizsa différentes ont été utilisées dans cette étude. Il était demandé à Griffin de décrire combien d’angles étaient présents dans l’objet créé par les figures en « pac-man ». Note : La plupart des expérimentations dans la littérature non-humaine utilise une figure identique pour les tests. Encore une fois, aucun chiffre n’était présent sur les stimuli de Griffin. Crédit photographique : Dr Irene Pepperberg
Les couleurs utilisées étaient celles testées dans une étude antérieure afin de s’assurer que Griffin interpréterait correctement les descripteurs utilisés pour ces couleurs imprimées (Pepperberg et al 2008). Le violet et l’orange étaient ajustés au sein d’un intervalle approprié car ces couleurs peuvent être problématiques à identifier pour les Gris du Gabon (Pepperberg 1994 ; 2006). La capacité des psittacidés à voir dans le spectre ultraviolet peut provoquer une perception des couleurs quelque peu différente de celle des humains (Pepperberg y Nakayama 2016, Goldsmith y Butler 2005, Carvalho et al 2011, Bowmaker et al 1994).
Les résultats des tests montraient une haute précision (Table 1). Griffin avait raison dès le premier test de l’expérimentation, ce qui suggère qu’il a immédiatement transféré ses réponses – sans entraînement – d’objets tridimensionnels non masqués à des dessins de formes masquées ou imaginaire. Griffin a répondu de façon correcte sur la forme véritable des figures, malgré le fait que toutes soient partiellement masquées ou non réelles physiquement. La plupart des erreurs se sont produites pour les polygones et les triangles à un et trois angles, qu’il semblait confondre à la fois dans les tâches modales et amodales (Pepperberg y Nakayama 2016). Néanmoins, ces erreurs étaient globalement rares, et ne suggèrent pas qu’il se concentrait uniquement sur les angles à 60 degrés impliqués.
| Table 1. Résumé des résultats au test pour Griffin | ||
|---|---|---|
| Paramètre mesuré | Pourcentage de précision * | Nombre d’essais corrects |
| Exécution modale | 76 | 29/38 |
| Exécution amodale | 70 | 23/33 |
| *La chance est associée à une précision de 20% | ||
Conclusion
Dans l’expérience visuelle humaine, lorsqu’un objet est partiellement caché par un obstacle, nous ne percevons pas seulement les morceaux ou les fragments de cet objets. Les parties qui sont directement visibles suffisent habituellement à reconnaitre l’ensemble de l’objet. Les humains tendent également à voir des objets qui sont créés par des illusions, tels que les figures de Kanizsa. Un Gris du Gabon au moins, Griffin, répond de façon similaires et ces résultats – particulièrement son transfert de figures tridimensionnelles solides à des stimuli bidimensionnels imaginaires ou masqués, sans aucun entrainement – impliquent l’existence d’un concept sophistiqué de ce qu’est une forme. Bien qu’il soit peu probable que cette information en tant que telle soit directement applicable lors de l’organisation d’un enrichissement environmental , elle nous rappelle l’intelligence de ces oiseaux et le besoin de telles stimulations dans leur environnement.
Références
Références
Bowmaker JK, Heath LA, Das D, Hunt DM. Spectral sensitivity and opsin structure of avian rod and cone visual pigments. Invest Ophthalmol Vis Sci 35:1708, 1994.
Carvalho LS, Knott B, Berg ML, et al. Ultraviolet-sensitive vision in long-lived birds. Proc Royal Society B 278:107-114, 2011. Available at http://rspb.royalsocietypublishing.org/content/278/1702/107. Accessed July 28, 2016.
Fujita K. Seeing what is not there: Illusion, completion, and spatiotemporal boundary formation in comparative perspective. In: Wasserman EA, Zentall TR (eds). Comparative Cognition: Experimental Explorations of Animal Intelligence. New York, NY: Oxford University Press; 2006: 37, 42-50.
Fujita K. Perceptual completion in rhesus monkeys (Macaca mulatta) and pigeons (Columba livia). Perception and Psychophysics 63:115-125, 2001.
Funk MS. Development of object permanence in the New Zealand parakeet (Cyanoramphus auriceps). Animal Learning and Behavior 24:375-383, 1996.
Goldsmith T, Butler BK. Color vision of the budgerigar (Melopsittacus undulates): Hue matches, tetrachromacy, and intensity discrimination. J Comp Physiol 191:933-951, 2005. Available at http://www.2ndchance.info/tameparrot-Goldsmith2005.pdf. Accessed July 28, 2016.
Pepperberg IM. Addition by a Grey parrot (Psittacus erithacus), including absence of quantity. J Comp Psychol 120:1-11, 2006.
Pepperberg IM. The Alex Studies; Cognitive and Communicative Abilities of Grey Parrots. Cambridge, MA: Harvard; 1999.
Pepperberg IM. Evidence for numerical competence in an African Grey parrot (Psittacus erithacus). J Comp Psychol 106:295-305, 1994.
Pepperberg IM, Funk MS. Object permanence in four species of psittacine birds: An African Grey parrot (Psittacus erithacus), an Illiger mini maaw (Ara maracana), a parakeet (Melopsittacus undulatus), and a cockatiel (Nymphicus hollandicus). Animal Learning and Behavior 18:97-108, 1990
Pepperberg IM, Nakayama K. Robust representation of shape in a Grey parrot (Psittacus erithacus). Cognition 153:146-160, 2016.
Pepperberg IM, Vicinay J, Cavanagh P. The Muller-Lyer illusion is processed by a Grey parrot (Psittacus erithacus). Perception 37:765-781, 2008.
Plowright CMS, Reid S, Kilian T. Finding hidden food: Behavior on visible displacement tasks by mynahs (Gracula religiosa) and pigeons (Columba livia). J Comp Psychology 112:13-25, 1998.
Tvardíková K, Fuchs R. Tits use amodal completion in predator recognition: A field experiment. Animal Cognition 13:609-615, 2010.
Vallortigara G. The cognitive chicken: Visual and spatial cognition in a nonmammalian brain. In: Wasserman EA, Zentall TR (eds). Comparative Cognition: Experimental Explorations of Animal Intelligence. New York, NY: Oxford University Press; 2006: 53-70.