Key Points
- Cell entry of coronaviruses depends on binding of the viral spike (S) proteins to cellular receptors and on spike “S” protein priming by host cell proteases.
- For a host cell to be receptive to COVID-19, there must be homology (3D structural alignment) between the spike’s receptor binding domain and the receptor binding site on the host cell.
- The novel coronavirus, COVID-19 or SARS-CoV-2, binds to the SARS-CoV receptor angiotensin-converting enzyme 2 (ACE2).
- COVID-19 requires the presence of the serine protease TMPRSS2 in the host to activate fusion of the viral envelope with the host cell. Once the virus has entered the host cell, viral replication begins.
- There are no published studies showing the expression and distribution of TMPRSS2 in ferret tissues. This data would allow us to identify the target organs in the ferret, as well as the real potential for pathogenicity of COVID-19 for this species.
- There is data evaluating the presence of TMPRSS2 in the tissues of dogs, cats, and pigs, which indicates that the ACE2 receptors of these species are unable to bind COVID-19.
- This addendum is intended to serve as a supplement to the information provided on coronaviruses and related public health concerns in “Coronaviruses in Animals: Implications for Veterinary Staff” by Dr. Thomas Donnelly.
One of the critical determinants of viral host range is the interaction between the coronavirus spike (S) protein (Fig 1) and the host cell receptor. Angiotensin-converting enzyme 2 (ACE2), a receptor for the attachment and uptake of the coronavirus that caused Severe Acute Respiratory Syndrome (SARS-CoV-1), is widely expressed with conserved primary structures throughout the animal kingdom, from fish, amphibians, and reptiles, to birds and mammals. Structural analysis also suggests that ACE2 from these animals can potentially bind to the receptor binding domain (RBD) of COVID-19’s spike protein (Fig 2). By itself, the structural analysis suggests these animals would all be possible natural hosts for the virus.
Fortunately, in order for a coronavirus to bind to a host cell releasing their genomic material inside it and starting the infection of the host, two conditions must be met. First, there must be homology between the receptor site of the host cell’s receptor, ACE2, and the RBD on the spike protein of the viral envelope. Additionally, a specific endogenous protease must be present on the host cell that is capable of “activating” the fusion between the RBD and ACE2. This second mechanism, which highlights the need for specific proteases for viral activation, greatly reduces the spectrum of “permissive” hosts to a specific virus, even if there are strong receptor analogies. Based on which and how much protease is present in a tissue, the infection can vary in severity, and the virus may have a greater or lesser virulence.
Understanding this concept is essential, as it explains why, in different animal species, a virus can have different organ tropism, but above all it can induce milder or more serious clinical signs.

Figure 1. Coronaviruses possess crown-like spike (S) proteins on their surface. Click image to enlarge.

Figure 2. In order for a host cell to be receptive to COVID-19, there must be homology between the receptor binding domain (located on the spike of the viral envelope) and the host cell’s angiotensin-converting enzyme 2 receptor or the receptor binding site. Photo credit: ScientificAnimations.com/C&EN. Click image to enlarge.
The peculiar structure of the spike “S” in COVID-19
“The spike is the business part as far as viral entry is concerned…It is in charge not only of attachment at the host cell surface, but also of fusing the viral and host cell membranes to allow the infection to start…”– David Veesler 1
A specific endogenous protease cleaves the S protein of COVID-19 into two subunits: the S1 and S2 subunits (Fig 3). A small RBD on the S1 subunit (amino acids 318–510) can then combine with the host cell receptor, ACE2. The entire receptor-binding loop, known as the receptor-binding motif (RBM) (amino acids 424–494), is located on the RBD and is responsible for complete contact with ACE2.

Figure 3. A small receptor binding domain on the S1 subunit of the spike (S) protein binds with the host cell receptor, angiotensin-converting enzyme 2. The receptor binding domain displays a concave surface during this interaction with the receptor. Photo credit: Bio Connect.com.
Glycosylation can also affect the interaction of the RBD with ACE2. N-glycosylation represents the attachment of a polysaccharide or glycan to a nitrogen atom or the amide nitrogen of an asparagine (Asn) residue of a protein. This type of binding is important for both the structure and function of virus spike and host cell receptor binding site (RBS) proteins. Among the 23 glycosylation sites on spike glycoprotein, two are in the RBD. In contrast with other coronaviruses, in COVID-19 an adjunctive glycosylation has been detected on asparagine Asn330 2, but not in N357. In addition, these N-linked glycosylation sites are fundamental not only for the ability of coronavirus to infect a novel species but also as a site for the attack of neutralizing antibodies. This is demonstrated in the case of the highly specific S230 antibody for COVID-19 (Fig 4).3 This type of binding is important both for the structure and for the function of virus spike and host-cell RBS protein N-glycosylation is acquired in the endoplasmic reticulum and Golgi apparatus during the intracellular phases of virus replication (Fig 5).

Figure 4. Schematic of complex glycans in the interplay between virus and host. Photo credit: SemanticScholar.org. Click image to enlarge.

Figure 5. N-glycosylation is acquired in the endoplasmic reticulum and Golgi apparatus during the intracellular phases of virus replication. Photo credit: ScienceDirect.com. Click image to enlarge.
The presence and role of the host cell specific endogenous protease
Many human respiratory viruses rely on interactions with host cell proteases for their activation, and this activation is necessary for their successful replication within host cells. Type II transmembrane protease, serine S1 member 2 (TMPRSS2 or epitheliasin) competently activates coronaviruses, influenza A, and other viruses, including members of Paramyxoviridae (Fig 6) Epitheliasin or TMPRSS2 is present in human airway epithelium as well as other organs and in the corresponding structures of different animal species.4
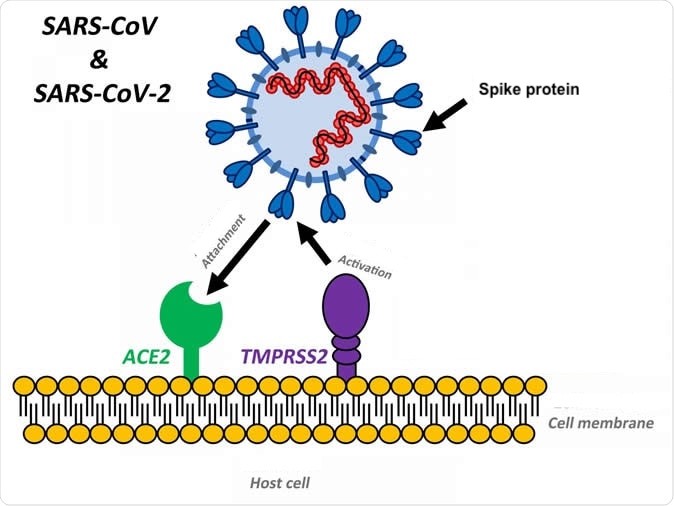
Figure 6. In addition to the host cell receptor angiotensin-converting enzyme 2 (ACE2), the host tissue must express a specific endogenous protease (TMPRSS2) that activates fusion of the virus receptor binding domain with the host cell membrane receptor binding site. Photo credit: SoundHealthandLastingWealth.com. Click image to enlarge.
Moreover, as polymorphisms in the TMPRSS2 gene were found to be associated with influenza severity in human patients (Cheng et al 2015), it appears that different influenza viruses might also take advantage of TMPRSS2 for spread in the human host. Non-human hosts, such as ferrets, swine, bats and probably others, could show a different receptivity to the COVID-19 virus, and different organ pathologies.
In order for a host to be receptive to COVID-19, there must be a homology between the spike’s RBD and the ACE2 receptor. In addition, the host tissue must express the specific serine protease TMPRSS2 to activate the fusion of the virus with the host cell.
Coronavirus infections are initiated by spike (S) protein activation by TMPRSS2 attachment to cell surface receptors by RBD, and fusion of the viral lipid envelope and endosomal membranes upon endocytosis to release the virus genome into the cytosol (Fig 7).
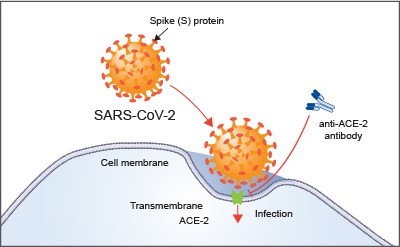
Figure 7. Angiotensin-converting enzyme 2 (ACE-2) is the host cell receptor responsible for mediating infection by SARS-CoV-2, the novel coronavirus responsible for coronavirus disease 2019 (COVID-19). Treatment with anti-ACE-2 antibodies disrupts the interaction between virus and receptor. Photo credit: RNDSystems.com
COVID-19 or SARS-CoV-2 uses the receptor ACE2 for entry and the serine protease TMPRSS2 for S protein priming. Protein priming is a generalized mechanism to initiate RNA synthesis of the virus within the host cell.
Some coronaviruses, like the SARS coronavirus of 2003 and the novel coronavirus COVID-19, are activated by TMPRSS2 and can thus be inhibited by TMPRSS2 inhibitors.5 A TMPRSS2 inhibitor approved for clinical use blocked entry and might constitute a treatment option (Fig 8).5

Figure 8. The novel coronavirus COVID-19 is activated by TMPRSS2 and can be inhibited by TMPRRSS2 inhibitors. Photo credit: ScienceDirect.com. Click image to enlarge.
What is the role of animals in the receptivity of COVID-19?
As described above, a high degree of homology between the viral spike protein and the host ACE2 receptor is not enough to allow the virus to enter the host cell. The serine protease TMPRSS2 must be simultaneously expressed on the cell to activate the fusion of viral cell membranes with the host cell wall.
FERRETS
To our knowledge, to date there are no published studies showing the expression and distribution of TMPRSS2 in ferret tissues. This data would allow us to identify the target organs in the ferret, as well as the real potential for pathogenicity of COVID-19 for this species.
Instead, we have data regarding the presence of TMPRSS2 in the tissues of the dog, cat, and pig, which indicates that the ACE2 receptors of these species are unable to bind COVID-19.
RODENTS
As with SARS, mice and rats are likely resistant to infection.
BIRDS
Birds are not believed to be susceptible to infection with COVID-19.
DOGS
Canine respiratory coronavirus (CRCoV) was first identified in 2003 in samples obtained from the respiratory tracts of dogs with canine infectious respiratory disease (CIRD), also known as kennel cough).6 Szczepanski et al demonstrated that infection sustained by CRCoV is characterized by a preliminary endocytic internalization, prior to membrane fusion, a process that requires caveolin-1 and dynamin but not TMPRSS2, which is missing in canine cells and tissues.6 Furthermore, in this species, fusion of viral and cellular membranes occurs before the endosome progresses to the late phase, indicating that the dog is not a permissive host for COVID-19.
CATS
Feline coronavirus (FCoV) (Alphacoronavirus 1 genus Alphacoronaivirus) 7, is divided into serotypes I and II based on the amino acid sequence of the spike (S) protein.8,9 Type I FCoV is the dominant serotype (80–90%) in Europe and Asia, and can also be divided into two biotypes, feline infectious peritonitis virus (FIP virus, FIPV; virulent FCoV) and feline enteric coronavirus (FECV; avirulent FCoV), according to its pathogenesis in animals.10
Feline coronavirus cell surface attachment and entry is not mediated by TMPRSS2, but through feline aminopeptidase N (fAPN), which acts as the RBS for viral infections. Feline coronaviuruses enter the cell through endocytosis after attachment to the feline aminopeptidase-N (fAPN).11 It is speculated that low endosomal pH triggers the conformation change of the spike protein of FCoV that releases the virus from the endosome to the cytosol. These studies are important in helping to exclude the domestic cat and others felids as permissive hosts to COVID-19 12, but this type of characterization regarding the receptor and its protease has never been performed in other wild felids. Therefore it cannot be excluded that the recent episodes of infection occurring in some tigers of the Wildlife Conservation Society’s Bronx Zoo, may be linked to a different receptor structure in these big felids, which were also sensitive to SARS and influenza viruses.13
PIGS
Porcine epidemic diarrhea virus (PEDV) and porcine deltacoronavirus (PDCoV) are important infectious disease concerns in pigs. These are alpha and delta coronaviruses, respectively.
Porcine TMPRSS2 is expressed through out the entire respiratory tract.14 In addition, TMPRSS2 protein expression has been identified in the digestive tract, some lymphoid organs, kidneys, and reproductive system. Using immunohistochemistry and reverse transcription PCR assays to evaluate TMPRSS2 mRNA expression, the author showed some interesting cell type localization in this species’ tissues.
Pigs are important natural hosts of influenza A viruses, and due to their susceptibility to swine, avian, and human viruses, they may serve as intermediate hosts supporting adaptation and genetic reassortment. Cleavage of the influenza virus surface (HA) by host cell proteases is essential for viral infectivity. Most influenza viruses, including human and swine viruses, are activated at a monobasic glycoprotein hemagglutinin (HA) cleavage site, in association with TMPRSS2 cleavage site. Peitsch et al showed that swine proteases homologous to TMPRSS2 and HAT, designated swTMPRSS2 and swAT, respectively, were expressed in several parts of the porcine respiratory tract, but also in different organs.15
Despite the expression of the TMPRSS2 protease in the swine respiratory tract, small changes or replacements of amino acids cause the spike protein of COVID-19 to not bind well to the cellular receptor of the pig. Therefore even though TMPRSS2 is well distributed in the pig, this species is not a good host for COVID-19 for this reason.7
Also view “Susceptibility of ferrets, cats, dogs, and different domestic animals to SARS-coronavirus-2”, a preliminary report that has not been peer-reviewed, which reports that SARS-CoV-2 replicates poorly in dogs, pigs, chickens, and ducks, but efficiently in ferrets and cats.*
References
References
1. Gray L. COVID-19 coronavirus spike holds infectivity details. UW Medicine Newsroom. Available at newsroom.uw.edu/news/covid-19-coronavirus-spike-holds-infectivity-details. Accessed April 1, 2020.
2. Wrapp D, Wang N, Corbett KS. et al. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 367(6483):1260-1263, 2020. doi: 10.1126/science.abb2507.
3. Tian X, Lic C, Huang A, et al. Potent binding of 2019 novel coronavirus spike protein by a SARS coronavirus-specific human monoclonal antibody. Emerg Microbes Infect. 9(1):382-385, 2020. doi: 10.1080/22221751.2020.
4. Tarnow C, Engels G, Arendt A, et al. TMPRSS2 is a host factor that is essential for pneumotropism and pathogenicity of H7N9 influenza A virus in mice. J Virol. 88(9):4744–4751, 2014. doi: 10.1128/JVI.03799-13.
Cheng Z, Zhou J, To KK, Chu H, Li C, Wang D, et al. (2015). Identification of TMPRSS2 as a susceptibility gene for severe 2009 Pandemic A(H1N1) Influenza and A(H7N9) Influenza. J Infect Dis.
5. Hoffmann M, Kleine-Weber H, Schroeder S, et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell. Mar 4. pii: S0092-8674(20)30229-4, 2020. doi: 10.1016/j.cell.2020.02.052. [Epub ahead of print]
6. Szczepanski A, Owczarek K, Milewska A, et al. Canine respiratory coronavirus employs caveolin-1-mediated pathway for internalization to HRT-18G cells. Vet Res. 49(1):55, 2018. doi: 10.1186/s13567-018-0551-9.
7. Su S, Wong G, Shi, W, et al. Epidemiology, genetic recombination, and pathogenesis of coronaviruses. Trends Microbiol. 24(6):490–502, 2016. doi: 10.1016/j.tim.2016.03.003.
8. Motokawa K, Hohdatsu T, Aizawa C, Koyama H, Hashimoto H. Molecular cloning and sequence determination of the peplomer protein gene of feline infectious peritonitis virus type I. Arch Virol. 140(3):469–480, 1995. doi: 10.1007/bf01718424.
9. Hohdatsu T, Okada S, Ishizuka Y, et al. The prevalence of types I and II feline coronavirus infections in cats. J Vet Med Sci. 54(3):557–562, 1992. doi: 10.1292/jvms.54.557.
10. Tekes G, Thiel HJ. Feline coronaviruses: pathogenesis of feline infectious peritonitis. Adv Virus Res. 96:193–218, 2016. doi: 10.1016/bs.aivir.2016.08.002.
11. Takano T, Wakayama Y, Doki T. Endocytic pathway of feline coronavirus for cell entry: differences in serotype-dependent viral entry pathway. Pathogens. 2019, 8(4): pii:E300, 2019. doi: 10.3390/pathogens8040300.
12. Gillespie EJ, Ho CLC, Balaji K, et al. Selective inhibitor of endosomal trafficking pathways exploited by multiple toxins and viruses. Proc Natl Acad Sci USA. 110(50):E4904–E4912, 2013. doi: 10.1073/pnas.1302334110.
13. United States Department of Agriculture. USDA statement on the confirmation of COVID-19 in a tiger in New York. USDA Web site. Apr 5, 2020. Available at aphis.usda.gov/aphis/newsroom/news/sa_by_date/sa-2020/ny-zoo-covid-19. Accessed April 5, 2020.
14. Owens JL. Immunohistochemical characterization of protease TMPRSS2 expression in porcine tissue. FASEB J. 33(1). Abstract Number:38.2. Published Online:1 Apr 2019. doi: 10.1096/fasebj.2019.33.1_supplement.38.2.
15. Peitsch C, Klenk HD, Garten W, Böttcher-Frietbertshäuser E. Activation of influenza A viruses by host proteases from swine airway epithelium. J Virol. 88(1):282-91, 2014. doi: 10.1128/JVI.01635-13.
Further reading
Bertram S, Heurich A, Lavender H, et al. Influenza and SARS-coronavirus activating proteases TMPRSS2 and HAT are expressed at multiple sites in human respiratory and gastrointestinal tracts. PLoS One. 7(4):e35876, 2012. doi: 10.1371/journal.pone.0035876.
Cheng Z, Zhou J, To KK, et al. Identification of TMPRSS2 as a susceptibility gene for severe 2009 pandemic A(H1N1) influenza and A (H7N9) influenza. J Infect Dis. 212(8):1214-21, 2015. doi: 10.1093/infdis/jiv246.
Mettelman RC, O’Brien A, Whittaker GR, Baker SC. Generating and evaluating type I interferon receptor-deficient and feline TMPRSS2-expressing cells for propagating serotype I feline infectious peritonitis virus. Virology. 537:226-236, 2019. doi: 10.1016/j.virol.2019.08.030.
Zhou P, Yang X, Wang X, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579(7798):270–273, 2020. doi: 10.1038/s41586-020-2012-7. Epub 2020 Feb 3.
Rossi G. Determinants of viral infection: an addendum to coronaviruses in animals. April 3, 2020. LafeberVet Web site. Available at https://lafeber.com/vet/determinants-of-viral-infection-an-addendum-to-coronaviruses-in-animals/